Every few months we highlight a subset of the new plasmids and viral preps in the repository through our hot plasmids articles. These articles provide brief summaries of recent plasmid deposits and we hope they'll make it easier for you to find and use the plasmids you need. If you'd ever like to write about a recent plasmid deposit please sign up here.

Here's what you'll find in this post:
- Anti-FLAG Frankenbodies for modular imaging of cellular targets
- Optimized genetic screening technique using Cas12a for multiplexed gene perturbation
- The Penn State Protein Ladder
- Engineered miniature CRISPR-Cas system for mammalian genome regulation and editing
Anti-FLAG Frankenbodies for modular imaging of cellular targets
by: Mike Lacy
Adding to their set of anti-HA frankenbodies (Zhao et al. 2019), Tim Stasevich and Hiroshi Kimura’s labs have recently published a set of anti-FLAG frankenbodies, a flexible, modular tool for labeling. Here, Liu et al. engineer a chimeric single chain variable fragment (scFv), grafting the binding region of a FLAG-binding scFv onto a well-folded scFv scaffold to improve its stability in cells, and use it in a variety of applications. This set of new anti-FLAG frankebodies includes mEGFP, mRuby2, iRFP713, SNAP-tag, and HaloTag fusions, while the anti-HA frankenbodies include mEGFP, mCherry, and HaloTag.
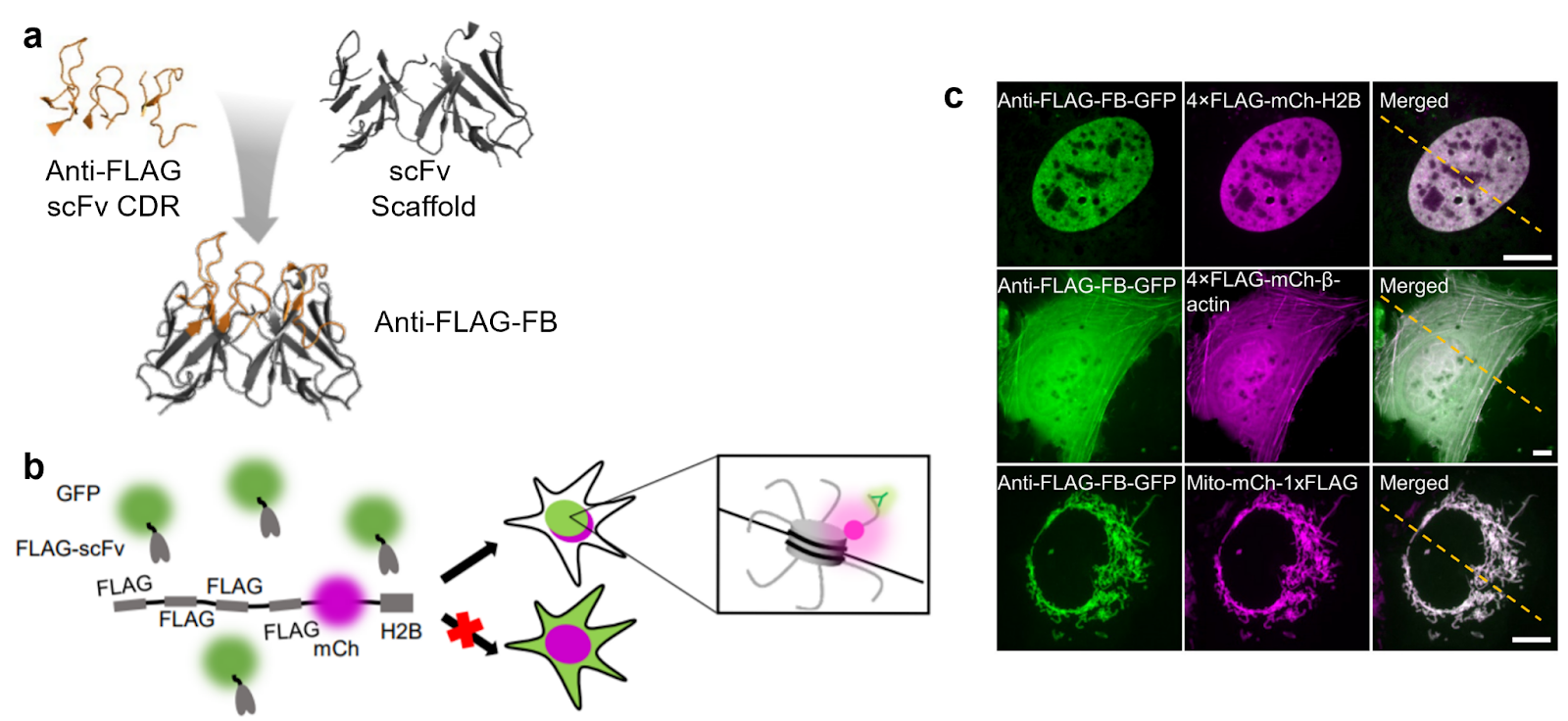 |
| Figure 1: (a) Anti-FLAG frankenbody (FB) construction from the complementarity determining region of an anti-FLAG scFv and a stable scFv scaffold. (b) Schematic of anti-FLAG-FB-GFP binding to FLAG-tag H2B for nuclear localization. (c) Applications of anti-FLAG-FB-GFP labeling various targets with nuclear (top, 4x-FLAG-mCherry-H2B), cytoskeletal (middle, 4xFLAG-mCherry-β-actin), and mitochondrial (mito-mCherry-1xFLAG) localization. Adapted from Liu et al. 2021. |
Anti-FLAG and anti-HA frankenbodies have a variety of applications, including live-cell imaging, single-molecule tracking, immunolabeling of fixed cells, Western blotting, and more. Given the popularity of FLAG and HA tags, you may not even need to clone new constructs to label your favorite protein! While the target only requires a small epitope tag (FLAG is eight amino acids, HA is nine), multiple repeats of the tag can recruit multiple frankenbodies to amplify the signal.

Liu et al. Genes Cells. 2021. https://doi.org/10.1111/gtc.12893,
Zhao et al. Nat Comm. 2019. https://doi.org/10.1038/s41467-019-10846-1.
Optimized genetic screening technique using Cas12a for multiplexed gene perturbation
by: Gabrielle Clouse
While many CRISPR-based genetic screening methods have primarily used Cas9 to knock out genes, researchers at the Genetic Pertubation Platform at the Broad Institute have expanded the genetic screening toolkit by optimizing a Cas12a-based method for screens. Cas12a’s ability to perform multiplex editing within a single expression cassette is advantageous to Cas9’s larger expression requirements (Figure 1), but Cas12a has generally been underutilized in genetic screens because of its lowered editing rate. This research aimed to increase the potential of Cas12a genetic screens by optimizing enzyme activity using an enhanced Cas12a variant (enAsCas12a), minimizing off-target effects by identifying guide selection rules, and reducing recombination by finding acceptable variations in DR sequences.
 |
| Figure 2: comparison of Cas 9 vs. Cas 12a expression cassettes when encoding multiple guides. While two guides require two arrays for Cas9, two guides can be streamlined into a single array for Cas12a. Image from DeWeirdt, et al. 2021 |
To validate this optimized genetic screening technique, researchers identified a synthetic lethal interaction between MARCH5 and WSB2 genes using Cas12a’s ability to knock out multiple genes in parallel. Lastly, this Cas12a-based screening method was used to create a genome-wide library capable of performing similarly to Cas9 screens in a compacted library. Overall, Cas12a’s ability to target multiple genes within a single cassette provides increased simplicity for combinatorial genetic screens, and this optimized toolkit furthers Cas12a’s potential in future genetic screens.

DeWeirdt PC, et al. Nat Biotechnol. 2021. doi: 10.1038/s41587-020-0600-6.
The Penn State Protein Ladder
by: Rachel Leeson
This simple, inexpensive plasmid-based ladder system is a cost-effective alternative to commercial ladders which also provides a useful yet low-pressure teaching opportunity for lab members or students new to protein expression and purification.
 |
| Figure 3: The Penn State Ladder run on SDS-Page gels. A: individually expressed proteins run in separate lanes. B: individually expressed and coexpressed proteins run as ladders. Image from Santilli, et al. 2021 |
Plasmids are used to express 10, 15, 20, 30, 40, 50, 60, 80 and 100 kD proteins in E. coli, either individually (~20,000 lanes/50 mLs) or through two polycistronic expression vectors (3570 lanes/50 mLs) coexpressing a set of either 10, 30, 50, and 100 kD proteins or 20, 40, 60, and 80 kD proteins. Plasmids containing 150 or 250 kD proteins are also available separately. The proteins, which are purified via histidine tags, are optimized for SDS-PAGE gels and can be detected on Western blots via engineered immunoglobulin binding domains.

Santilli, R.T., et al. Sci Rep (2021). https://doi.org/10.1038/s41598-021-96051-x
Engineered miniature CRISPR-Cas system for mammalian genome regulation and editing
by: Rachel Leeson
The large size of CRISPR-Cas effectors and fusion proteins, such as Cas9 and Cas12a, creates challenges for in vivo delivery and cell engineering applications. CasMINI, a new miniature CRISPR-Cas system for editing and gene activation, is composed of only 529 amino acids, less than half the size of either Cas9 or Cas12a. Despite its small size, CasMINI is comparable in efficacy to dCas12a systems in both gene activation and base editing applications.
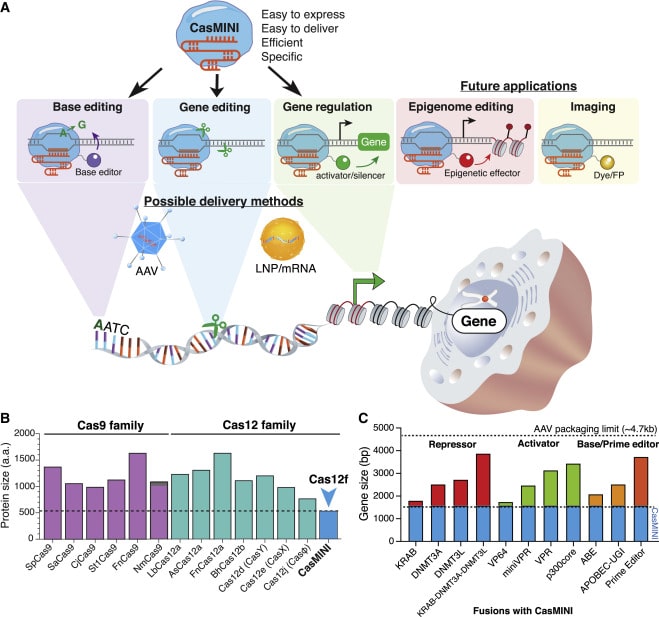 |
| Figure 4: Overview of CasMINI for genome engineering applications. Image from Xiaoshu, et. al. 2021. |
Created by the fusion of Cas12f with a transcriptional activator, CasMINI is able to induce high levels of gene activation for both reporter and endogenous genes. It is highly specific in mammalian cells without significant off-target effects, yet small enough to be packaged in AAV. A dCasMini-mediated adenine base editor allows for base editing, while gene editing can be done via a nuclease-active CasMINI. This small yet powerful system opens up new possibilities in both cell engineering and gene therapy.

Xiaoshu X, et al. Molecular Cell. 2021. https://doi.org/10.1016/j.molcel.2021.08.008.

Topics: Hot Plasmids





Leave a Comment