This post was contributed by Alessia Armezzani, scientific communication manager at genOway.
A few decades ago, the brain remained elusive, not from a lack of intellectual curiosity on the part of scientists but, rather, from the limited technologies available. Over the past few years, however, remarkable technological advances have taken researchers to the threshold of a revolution in modern neuroscience, an era in which technologies like FLEx and optogenetics meet, allowing scientists to investigate the fundamental basis of both brain function and dysfunction.
First, let’s start with the basics: What is optogenetics?
Optogenetics: Controlling neurons with light
In 2005, a team from Stanford University, led by neurobiologist Karl Deisseroth, combined optical and genetic methods to photo-stimulate mammalian neurons and alter synaptic events: a new technology was born, and the term “optogenetics” was coined (Boyden et al., 2005; Deisseroth et al., 2006).
Despite its relatively recent development, the history of optogenetics begins almost 50 years ago with the discovery of microbial rhodopsins, light-sensitive proteins that absorb photons for energy conversion or the induction of intra- or intercellular signaling pathways (Ernst et al., 2014). Microbial rhodopsins are encoded by opsin genes, and include bacteriorhodopsins, halorhodopsins, and channelrhodopsins, the first two found in the Archaea, the third in green algae (Oesterhelt and Stoeckenius, 1971; Matsuno-Yagi and Mukohata, 1977; Foster et al., 1984; Harz and Hegemann, 1991). These proteins control the flow of electric charges across cell membranes and maintain membrane potential in response to visible light (Zhang et al., 2011).
What do microbial rhodopsins and neurons have in common? Neuronal stimulations are triggered by the movement of ions across the axon's membrane: once a certain number of positive ions crosses the cell membrane, a threshold is reached and the neuron fires, sending the electrical signal down the axon (Lodish et al., 2000). Most neurons in the brain are not naturally light-sensitive, so the selective expression of opsin genes in targeted cells makes it possible to control neuronal activity with a specificity far greater than the one that can be achieved using pharmacological or electrophysiological methods (Deisseroth, 2011; Mei and Zhang, 2012; Towne et al., 2016).
Optogenetic neuromodulation is made possible by the light: when hit with the correct wavelength, microbial rhodopsins enable ions to flow across the axon’s membrane, thereby controlling neural activity (Bernstein and Boyden, 2011; Mei and Zhang, 2012). For example, blue light activates channelrhodopsins that, in turn, trigger neural excitation, whereas yellow light activates halorhodopsins to silence neuronal activity (Figure 1) (Wiegert et al., 2017).
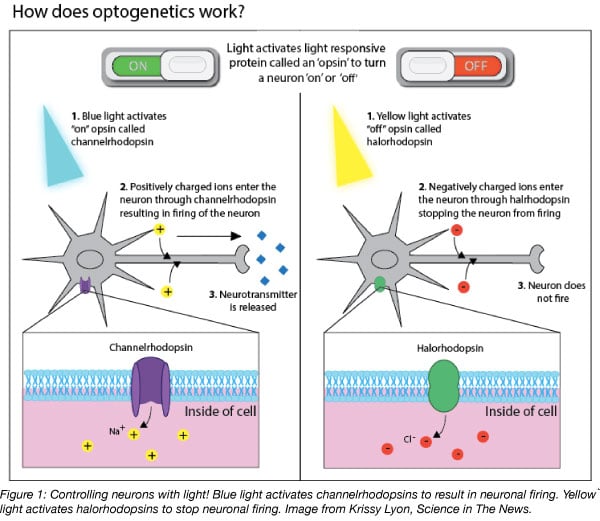
How to get an opsin into the brain
In order to express opsins within the brain, researchers inject genetically modified viruses encoding microbial rhodopsins into specific cerebral regions. The resulting viral-infected neurons are subsequently photo-stimulated through fiberoptic cannulas directly implanted in the injection site and connected to a laser. The laser flashes light of specific wavelengths, selectively turning neuron activity on or off (Atasoy et al., 2008; Taylor et al., 2016; Hooper and Maguire, 2016).
Recombinant adeno-associated viruses (rAAVs) have become increasingly popular for gene delivery in the central nervous system due to their relatively stable extra-chromosomally long-term expression and low ability to induce innate immune responses (Fenno et al., 2011). However, rAAVs lack the ability to specifically infect defined neuronal subpopulations, and this represents a major downside in optogenetics (Belzung et al., 2014). This could be overcome by fusing opsin genes to cell-type specific promoters; however, those usually drive weak expression of downstream genes. Strong cell-specific promoters, on the other side, are too long for rAAVs, which can only package sequences shorter than 5 kb (Hirsch et al., 2016; Hudry and Vandenberghe, 2019). How to achieve strong opsin expression in specific neuronal cell types then? This is where FLEx enters the scene.
FLEx: A light when night is about you!
The FLEx (for flip-excision) switch (aka DIO (Double-floxed Inverse Orientation) or DO (Double-floxed Orientation)) is a powerful tool that provides precise temporal and spatial control of gene expression in vivo (Schnütgen et al., 2003). This is achieved through site-specific recombinases such as Cre or Flp that induce DNA recombination at defined recognition sites (i.e., loxP and FRT, respectively) (Abremski and Hoess, 1984; Van Duyne, 2001; Christenson Wick and Krook-Magnuson, 2018).
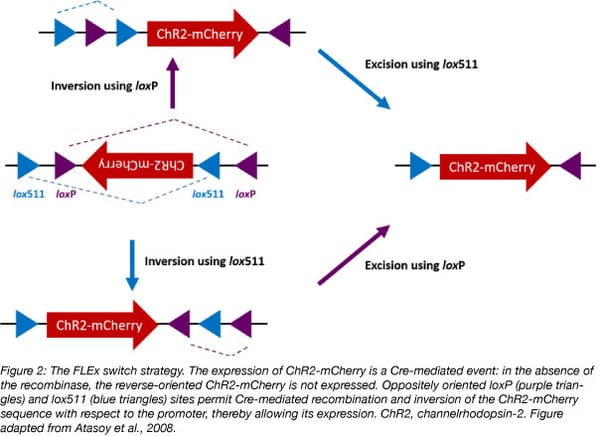
Optogenetic FLEx vectors contain a strong promoter upstream of an opsin gene (e.g., channelrhodopsin-2, ChR2) fused to a reporter gene (e.g., mCherry) to easily detect opsin-expressing cells. The resulting fusion gene is inserted in the antisense orientation with respect to the promoter to prevent its expression, and is flanked (“floxed”) by two sets of incompatible recognition sites (e.g., loxP and lox511) in opposite orientations. Since Cre does not cause recombination between mismatched recognition sites, its presence induces first opsin inversion, and then lox sites excision, therefore locking the opsin into the correct orientation to be transcribed (Figure 2) (Sharma and Zhu, 2014).
How does FLEx enable strong opsin expression in specific neuronal cells? This can be achieved using Cre-dependent viruses, such as the optogenetic FLEx vectors in combination with transgenic animals or rAAVs expressing Cre under a specific cell-type promoter. Once injected into the brain, the viruses infect all the cells with an inactive opsin gene; in Cre expressing cells, however, Cre induces the recombination of the double-floxed opsin construct, thereby enabling its expression under a strong promoter only in specific neuronal cells. FLEx ensures, therefore, spatial accuracy and strong opsin expression, both of which are essential in optogenetics to study physiological and behavioral processes (Abdallah et al., 2018; Deubner et al., 2019).
For example, Taylor and colleagues used rAAV FLEx-based vectors to study the neural circuitry behind the mechanisms of sleep and anesthesia. In Cre transgenic mice, they targeted the dopamine (DA) neurons of the ventral tegmental area (VTA) of the brain. In particular, they found that optogenetic stimulations of the DA neurons produce behavioral and electroencephalography evidences of arousal in mice previously subjected to steady-state general anesthesia (Taylor et al., 2016).
These very same neurons also play a central role in motivated behaviors (Juarez and Han, 2016). Not so long ago, however, it was completely unclear which subpopulation of DA neurons could activate appetitive rather than aversive stimuli. In a recent paper, de Jong and colleagues used rAAV FLEx-based vectors to answer this question, mapping and characterizing the activity of the DA neurons of the VTA. Using in vivo optogenetic stimulations, they simultaneously recorded the electrical impulses of discrete subpopulations of this brain area, demonstrating that it is possible to separate neuronal inputs to induce aversion- or reward-related behaviors. The high spatio-temporal precision and reversible modulation of FLEx vectors combined with the use of several Cre transgenic mice targeting different brain areas enabled the drawing of a detailed and functional topography of the neural circuit architecture of the brain regions associated with motivated behaviors (de Jong et al., 2019). FLEx vectors represent, therefore, an ideal partner to optogenetics to understand the cellular and molecular mechanisms of the brain in vivo.

Many thanks to our guest blogger Alessia Armezzani from genOway.
 Alessia Armezzani received her PhD in Virology from the University of Glasgow. She then moved to France where she worked as a post-doctoral fellow on developmental biology at the École Normale Supérieure de Lyon. Her love for educational science led her to pursue a teaching career, and she now is a very happy scientific communication manager at genOway.
Alessia Armezzani received her PhD in Virology from the University of Glasgow. She then moved to France where she worked as a post-doctoral fellow on developmental biology at the École Normale Supérieure de Lyon. Her love for educational science led her to pursue a teaching career, and she now is a very happy scientific communication manager at genOway.
Acknowledgement:
Through a partnership with genOway, we are able to distribute materials containing FLEx technology. Read the genOway press release for more information.
References
Abdallah, Khaled, et al. "Adeno-associated virus 2/9 delivery of Cre recombinase in mouse primary afferents." Scientific reports 8.1 (2018): 7321. PubMed PMID: 29743652. PubMed Central PMCID: PMC5943452.
Abremski, Ken, and Ronald Hoess. "Bacteriophage P1 site-specific recombination. Purification and properties of the Cre recombinase protein." Journal of Biological Chemistry 259.3 (1984): 1509-1514. PubMed PMID: 6319400.
Atasoy, Deniz, et al. "A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping." Journal of Neuroscience 28.28 (2008): 7025-7030. PubMed PMID: 18614669.
Belzung, Catherine, Marc Turiault, and Guy Griebel. "Optogenetics to study the circuits of fear-and depression-like behaviors: a critical analysis." Pharmacology Biochemistry and Behavior 122 (2014): 144-157. PubMed PMID: 24727401.
Bernstein, Jacob G., and Edward S. Boyden. "Optogenetic tools for analyzing the neural circuits of behavior." Trends in cognitive sciences 15.12 (2011): 592-600. PubMed PMID: 22055387. PubMed Central PMCID: PMC3225502.
Boyden, Edward S., et al. "Millisecond-timescale, genetically targeted optical control of neural activity." Nature neuroscience 8.9 (2005): 1263. PubMed PMID: 16116447.
de Jong, Johannes W., et al. "A neural circuit mechanism for encoding aversive stimuli in the mesolimbic dopamine system." Neuron 101.1 (2019): 133-151. PubMed PMID: 30503173. PubMed Central PMCID: PMC6317997.
Deisseroth, Karl, et al. "Next-generation optical technologies for illuminating genetically targeted brain circuits." (2006): 10380-10386. PubMed PMID: 17035522. Pubmed Central PMCID: PMC2820367.
Deisseroth, Karl. "Optogenetics." Nature methods 8.1 (2011): 26. PubMed PMID: 21191368.
Deubner, Jan, Philippe Coulon, and Ilka Diester. "Optogenetic approaches to study the mammalian brain." Current opinion in structural biology 57 (2019): 157-163. PubMed PMID: 31082625.
Ernst, Oliver P., et al. "Microbial and animal rhodopsins: structures, functions, and molecular mechanisms." Chemical reviews 114.1 (2013): 126-163. PubMed PMID: 24364740. PubMed Central PMCID: PMC3979449.
Fenno, Lief, Ofer Yizhar, and Karl Deisseroth. "The development and application of optogenetics." Annual review of neuroscience 34 (2011). PubMed PMID: 21692661. PubMed Central PMCID: PMC6699620.
Foster, Kenneth W., et al. "A rhodopsin is the functional photoreceptor for phototaxis in the unicellular eukaryote Chlamydomonas." Nature 311.5988 (1984): 756. PubMed PMID: 6493336.
Harz, Hartmann, and Peter Hegemann. "Rhodopsin-regulated calcium currents in Chlamydomonas." Nature 351.6326 (1991): 489.
Hirsch, Matthew L., Sonya J. Wolf, and R. J. Samulski. "Delivering transgenic DNA exceeding the carrying capacity of AAV vectors." Gene Therapy for Neurological Disorders. Humana Press, New York, NY, 2016. 21-39. PubMed PMID: 26611576. PubMed Central PMCID: PMC4971574.
Hooper, Andrew, and Jamie Maguire. "Characterization of a novel subtype of hippocampal interneurons that express corticotropin‐releasing hormone." Hippocampus 26.1 (2016): 41-53. PubMed PMID: 26135556.
Hudry, Eloise, and Luk H. Vandenberghe. "Therapeutic AAV gene transfer to the nervous system: a clinical reality." Neuron 101.5 (2019): 839-862. PubMed PMID: 30844402.
Juarez, Barbara, and Ming-Hu Han. "Diversity of dopaminergic neural circuits in response to drug exposure." Neuropsychopharmacology 41.10 (2016): 2424. PubMed PMID: 26934955. PubMed Central PMCID: PMC4987841.
Lodish, Harvey et al. Molecular Cell Biology. New York: W. H. Freeman, 2000. Bookshelf ID: NBK21668.
Matsuno-Yagi, Akemi, and Yasuo Mukohata. "Two possible roles of bacteriorhodopsin; a comparative study of strains of Halobacterium halobium differing in pigmentation." Biochemical and biophysical research communications 78.1 (1977): 237-243. PubMed PMID: 20882.
Mei, Yuan, and Feng Zhang. "Molecular tools and approaches for optogenetics." Biological psychiatry 71.12 (2012): 1033-1038. PubMed PMID: 22480664.
Oesterhelt, Dieter, and Walther Stoeckenius. "Rhodopsin-like protein from the purple membrane of Halobacterium halobium." Nature new biology 233.39 (1971): 149. PubMed PMID: 4940442.
Schnütgen, Frank, et al. "A directional strategy for monitoring Cre-mediated recombination at the cellular level in the mouse." Nature biotechnology 21.5 (2003): 562. PubMed PMID: 12665802.
SSharma, Suveena, and Jinfang Zhu. "Immunologic applications of conditional gene modification technology in the mouse." Current protocols in immunology 105.1 (2014): 10-34. PubMed PMID: 24700321. PubMed Central PMCID: PMC4100247.
Taylor, Norman E., et al. "Optogenetic activation of dopamine neurons in the ventral tegmental area induces reanimation from general anesthesia." Proceedings of the National Academy of Sciences 113.45 (2016): 12826-12831. PubMed PMID: 27791160. PubMed Central PMCID: PMC5111696.
Towne, Chris, and Kimberly R. Thompson. "Overview on research and clinical applications of optogenetics." Current protocols in pharmacology 75.1 (2016): 11-19. PubMed PMID: 27960028.
Van Duyne, Gregory D. "A structural view of cre-loxp site-specific recombination." Annual review of biophysics and biomolecular structure 30.1 (2001): 87-104. PubMed PMID: 11340053.
Wick, Zoé Christenson, and Esther Krook-Magnuson. "Specificity, versatility, and continual development: the power of optogenetics for epilepsy research." Frontiers in cellular neuroscience 12 (2018). PubMed PMID: 29962936. PubMed Central PMCID: PMC6010559.
Wiegert, J. Simon, et al. "Silencing neurons: tools, applications, and experimental constraints." Neuron 95.3 (2017): 504-529. PubMed PMID: 28772120. PubMed Central PMCID: PMC5830081.
Zhang, Feng, et al. "The microbial opsin family of optogenetic tools." Cell 147.7 (2011): 1446-1457. PubMed PMID: 22196724. PubMed Central PMCID: PMC4166436.
Resources on the Addgene blog
- Learn more about FLEx vectors
- Download our Plasmids 101 eBook
- Read our plasmids blog posts
Resources on the Addgene website
- Visit our molecular biology reference
- Find Cre-lox plasmids at Addgene
- Browse plasmids at Addgene
Topics: Optogenetics, Other Plasmid Tools, Plasmids





Leave a Comment