Prior to the discovery of CRISPR/Cas systems, gene activation across multiple loci was an arduous process. When using zinc finger proteins or TALE proteins, proteins had to be re-engineered for each gene, making wide-scale gene activation seem next to impossible. The development of CRISPR/Cas systems, however, greatly improved the simplicity of gene activation: rather than requiring protein engineering for each loci, CRISPR/Cas systems only require changing the programmable guide RNA.
Gene activation by dCas9, also referred to as CRISPRa, was initially published in 2013 (Bikard et al., 2013, Perez-Pinera et al., 2013). In the years that followed, innovative methods greatly improved CRISPRa, expanding its practicality and popularity in research (Tanenbaum et al., 2014, Konermann et al., 2015, Chavez et al., 2015).
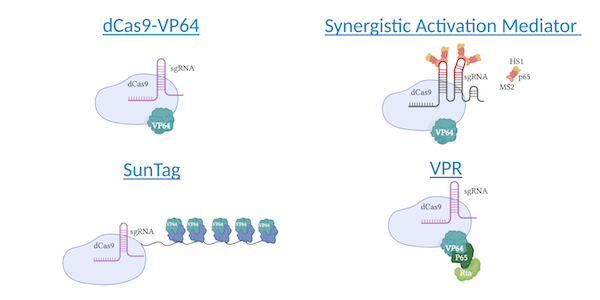
Most popular methods for CRISPR activation
CRISPR activation uses dCas9, a CRISPR protein variant lacking its endonuclease ability, to bind to genes without editing the genome (Qi et al., 2013). To target specific sequences, CRISPR/Cas systems rely on a guide RNA complementary to the sequence of interest. Upon binding, CRISPRa systems recruit transcription factors to increase gene expression. CRISPRa methods vary in their transcriptional activators: some methods rely on fusion proteins while others re-engineer components of Cas systems themselves. SunTag, SAM, and VPR have all shown significant improvements upon the initial dCas9-VP64 method, so there are multiple options to choose from when looking to activate genes across diverse cell lines.
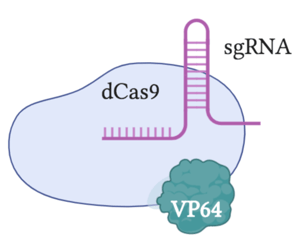
dCas9-VP64
Description
CRISPR activation can occur through fusing dCas9 with VP64, a strong transcriptional activation domain. Guided by dCas9, VP64 recruits transcriptional machinery to specific sequences, causing targeted gene regulation. This can be used to activate transcription during either initiation or elongation, depending on which sequence is targeted.
Performance
dCas9-VP64 activation is generally thought of as the “first generation” CRISPR activator. While it requires a relatively simple construct, it exhibits modest levels of gene activation, with some genes experiencing around 2-fold activation levels. Although other methods have been able to achieve much higher activation, dCas9-VP64 is great for experiments that requires modest gene activation.
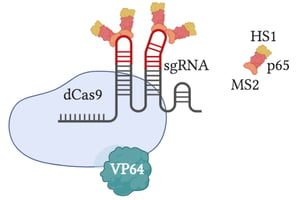
Synergistic Activation Mediator (SAM)
Description
SAM uses specially engineered sgRNAs to increase transcription. This is done through creating a dCas9/VP64 fusion protein engineered with aptamers that bind to MS2 proteins. These MS2 proteins then recruit additional activation domains (HS1 and p65).
Performance
When targeting single genes, SAM consistently shows the highest levels of gene activation compared to other CRISPR activators, making it a popular method for gene activation experiments. In cases of multiplex gene regulation (activating multiple genes at once), however, SAM exhibits activation levels comparable to other popular activation methods (VPR and SunTag) (Chavez et al., 2016).
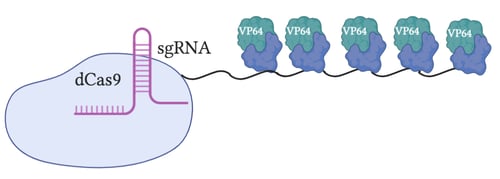
SunTag
Description
Rather than using a single copy of VP64 per each dCas9, SunTag uses a repeating peptide array to fused with multiple copies of VP64. By having multiple copies of VP64 at each loci of interest, this allows more transcriptional machinery to be recruited per targeted gene.
Performance
SunTag performs better than first generation activators while showing lower activation levels than SAM. One drawback of this method is its construction: it relies on antibody chains, which are relatively large and are not expressed consistently throughout cells.
VPR
Description
VPR fuses a tripartite complex with dCas9 to activate transcription. This complex consists of the VP64 activator used in other CRISPR activation methods, as well as two other potent transcriptional activators (p65 and Rta). These transcriptional activators work in tandem to recruit transcription factors.
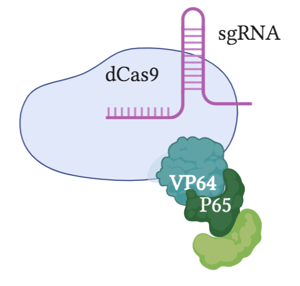
Performance
Generally, VPR is found to have significantly higher activation levels than the initial dCas9-VP64 activator, but lower levels compared to the SAM system. Its activation levels are similar to that of SunTag. An advantage to this method compared to other notable CRISPR activators is that it requires a fusion protein, rather than relying on a two-component system dependent on gRNA design (SAM) or peptide design (SunTag). This streamlines its delivery, making it a common choice for CRISPR activation.
Newer methods for CRISPR activation
While SAM, SunTag, and VPR have become the most popular methods for CRISPR activation, research in the area has continued to develop. These newer methods have not yet been rigorously compared to other CRISPR activators, but their initial results show promise for CRISPR-based gene activation.
dCas9-CBP
In this method, dCas9 is fused to CBP, an histone acetyltransferase domain capable of rearranging chromatin structure. This method has shown higher activation levels than SAM in some loci across Drosophila cell lines (Sajwan et al., 2019).
SPH
SPH combines components of SAM and SunTag activators, fusing the epitope tag of SunTag with the P65-HSF activation domains used in SAM. This method has been shown to activate genes at 2- to 3-fold higher than SAM, SunTag, and VPR (Zhou et al., 2018).
We hope this helps you chose a CRISPR activator for your experiment. Good luck with your experiment!

References
Bikard D, Jiang W, Samai P, Hochschild A, Zhang F, Marraffini LA (2013) Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Research 41:7429–7437 . https://doi.org/10.1093/nar/gkt520
Chavez A, Scheiman J, Vora S, Pruitt BW, Tuttle M, P R Iyer E, Lin S, Kiani S, Guzman CD, Wiegand DJ, Ter-Ovanesyan D, Braff JL, Davidsohn N, Housden BE, Perrimon N, Weiss R, Aach J, Collins JJ, Church GM (2015) Highly efficient Cas9-mediated transcriptional programming. Nat Methods 12:326–328 . https://doi.org/10.1038/nmeth.3312
Chavez A, Tuttle M, Pruitt BW, Ewen-Campen B, Chari R, Ter-Ovanesyan D, Haque SJ, Cecchi RJ, Kowal EJK, Buchthal J, Housden BE, Perrimon N, Collins JJ, Church G (2016) Comparison of Cas9 activators in multiple species. Nat Methods 13:563–567 . https://doi.org/10.1038/nmeth.3871
Konermann S, Brigham MD, Trevino AE, Joung J, Abudayyeh OO, Barcena C, Hsu PD, Habib N, Gootenberg JS, Nishimasu H, Nureki O, Zhang F (2014) Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 517:583–588 . https://doi.org/10.1038/nature14136
Perez-Pinera P, Kocak DD, Vockley CM, Adler AF, Kabadi AM, Polstein LR, Thakore PI, Glass KA, Ousterout DG, Leong KW, Guilak F, Crawford GE, Reddy TE, Gersbach CA (2013) RNA-guided gene activation by CRISPR-Cas9–based transcription factors. Nat Methods 10:973–976 . https://doi.org/10.1038/nmeth.2600
Qi LS, Larson MH, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, Lim WA (2013) Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 152:1173–1183 . https://doi.org/10.1016/j.cell.2013.02.022
Sajwan S, Mannervik M (2019) Gene activation by dCas9-CBP and the SAM system differ in target preference. Sci Rep 9: . https://doi.org/10.1038/s41598-019-54179-x
Tanenbaum ME, Gilbert LA, Qi LS, Weissman JS, Vale RD (2014) A Protein-Tagging System for Signal Amplification in Gene Expression and Fluorescence Imaging. Cell 159:635–646 . https://doi.org/10.1016/j.cell.2014.09.039
Zhou H, Liu J, Zhou C, Gao N, Rao Z, Li H, Hu X, Li C, Yao X, Shen X, Sun Y, Wei Y, Liu F, Ying W, Zhang J, Tang C, Zhang X, Xu H, Shi L, Cheng L, Huang P, Yang H (2018) In vivo simultaneous transcriptional activation of multiple genes in the brain using CRISPR–dCas9-activator transgenic mice. Nat Neurosci 21:440–446 . https://doi.org/10.1038/s41593-017-0060-6
Additional resources on the Addgene blog
- Browse our many CRISPR blog posts
- Get the basics of CRISPR in the CRISPR 101 blog series
Resources on Addene.org
Topics: CRISPR





Leave a Comment